Le Infezioni in Medicina, n. 1, 41-50, 2022
doi: 10.53854/liim-3001-5
REVIEWS
COVID-19-induced immune thrombocytopenic purpura; Immunopathogenesis and clinical implications
Mohammad Bahadoram1, Ali Saeedi-Boroujeni2, Mohammad-Reza Mahmoudian-Sani1, Helai Hussaini3, Shakiba Hassanzadeh1
1Thalassemia and Hemoglobinopathy Research Center, Health Research Institute, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran;
2Department of Immunology, Faculty of Medicine, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran;
3Department of Hematology, Faculty of Medicine, Kabul Medical University, Kabul, Afghanistan
Article received 12 December 2021, accepted 13 February 2022
Corresponding author
Shakiba Hassanzadeh
E-mail: shak.hassanzadeh@gmail.com
SummaRY
Following the outbreak of the COVID-19 pandemic, millions of people around the world have been affected with SARS-CoV-2 infection. In addition to the typical symptoms, thrombotic events, lymphopenia, and thrombocytopenia have been reported in COVID-19 patients. Immune thrombocytopenic purpura (ITP) is one of the thrombotic events that occur in some COVID-19 patients. Hyperinflammation, cytokine storms, and immune dysregulation in some patients are the cause to the main COVID-19 complications such as ALI (acute lung injury), acute respiratory distress syndrome (ARDS), and multiple organ failure. Disruption in the differentiation of T-cells, enhanced differentiation of Th17 and Th1, cell death (pyroptosis), hyper-inflammation and dysfunction of inflammatory neutrophils and macrophages, and hyperactivity of NLRP3-inflammasome are among the important factors that may be the cause to COVID-19-induced ITP. This study aimed to give an overview of the findings on the immunopathogenesis of ITP and COVID-19-induced ITP. Further studies are required to better understand the exact immunopathogenesis and effective treatments for ITP, especially in inflammatory disorders.
Keywords: COVID-19, SARS-CoV-2, inflammasome, immune thrombocytopenic purpura, platelets, immune dysregulation.
INTRODUCTION
Severe acute respiratory syndrome coronavirus (SARS-CoV-2) infection has been officially declared as a pandemic on March 11, 2020, and is still a major global threat. Although the main clinical manifestations of coronavirus disease 2019 (COVID-19) are respiratory, its range of clinical manifestations is extensive and may include neurological, gastrointestinal (GI), hepatic, and cardiac manifestations with varying degrees [1]. On the other hand, changes in laboratory parameters such as elevated liver enzymes, increased creatinine, electrolyte imbalance, a significant increase in C-reactive protein (CRP), and hyper-inflammation are very common in SARS-CoV-2 infection [2]. Various hematological disorders including thrombotic events, lymphopenia, thrombocytopenia, immune thrombocytopenia [or immune thrombocytopenic purpura (ITP)] have been reported in SARS-CoV-2 infection [3, 4]. ITP is defined as a platelet count of less than 100,000 per microliter with no evidence of leukopenia or anemia [5]. ITP is usually detected incidentally in patients with platelet counts above 50,000 per microliter. Excessive bruising with minor trauma occurs in those with platelet counts of 30,000 to 50,000 per microliter. In addition, patients with platelet counts between 10,000 and 30,000 per microliter spontaneously develop petechiae or ecchymoses, and those with counts less than 10,000 per microliter are at risk for internal bleeding [6].
ITP has a higher prevalence among children than adults. The annual prevalence of ITP in adults is 22 per million people [7-9]. The exact etiology of ITP is still unknown but genetic and other triggering factors appear to be involved [10, 11]. For example, some viral infections have been associated with secondary ITP such as human immunodeficiency virus (HIV), hepatitis C virus (HCV), cytomegalovirus (CMV), Epstein-Barr virus (EBV), and varicella-zoster virus (VZV) [8, 12]. It has been suggested that the viral infection-induced antibodies may cross-react with normal platelets and cause platelet destruction [13].
It is more prevalent in individuals older than 50 years. Most cases of COVID-19-induced ITP have occurred within 2-3 weeks of SARS-CoV-2 infection and had recovered in less than one week [4]. The patients have shown a good response to treatment, especially with intravenous immunoglobulin (IVIG) and/or corticosteroids [12, 14]. Alonso-Beato et al. emphasized that COVID-19-induced ITP may occur both in mild and severe COVID-19 and at any time during COVID-19’s course [12].
The diagnosis of ITP is established by ruling out other causes of thrombocytopenia [15]. It has two criteria:
1) The presence of thrombocytopenia with other normal findings (complete blood count and on peripheral blood slides);
2) The absence of other clinical features of medical conditions that cause secondary ITP such as systemic lupus erythematosus (SLE), antiphospholipid syndrome (APA), and chronic lymphoid leukaemia (CLL) [10].
This study aimed to give an overview of the immunopathogenesis and clinical implications of COVID-19-induced ITP.
METHODS
We searched PubMed/Medline, Web of Science, Scopus, and Google Scholar databases using the following keywords: Immune thrombocytopenic purpura; COVID-19; COVID-19-associated-ITP; ITP; COVID-19-induced ITP; COVID-19; SARS-CoV-2; Inflammasome; Platelets; Immune dysregulation.
COVID-19-induced Immune Thrombocytopenic Purpura
COVID-19-induced ITP has been reported to cause more severe thrombocytopenia, significantly lower platelet counts, more bleeding episodes, and more intracranial hemorrhages (ICHs) compared to non-COVID-19-induced ITP [12]. The incidence of thrombocytopenia in COVID-19 patients has been variable across studies. In April 2020, one of the first case reports of ITP in a COVID-19 patient was reported [16]. Moreover, mild thrombocytopenia has been observed in up to one-third of these patients with higher rates in patients with severe COVID-19 (57.7%) compared with non-severe COVID-19 (31.6%) [17]. A study reported that the majority (71%) of the cases of COVID-19-associated ITP were elderly (age >50 years). They also reported that 75% of the cases had moderate-to-severe COVID-19 and about 30% to 40% were asymptomatic [3]. A recent systematic review and meta-analysis reported that most of the patients that had developed COVID-19-associated ITP were male (54.8%) and over 50 years of age (median age of 63 years). In addition, most of the patients had developed ITP within 2-3 weeks after SARS-CoV-2 infection and recovered within a week [4].
Clinical presentations
Although ITP may manifest acutely and suddenly, in most cases it has an insidious onset. Bleeding in symptomatic patients with ITP can range from mild petechiae and bruising to severe haemorrhage. Thrombocytopenia-induced bleeding often occurs as mucocutaneous bleeding [15]. Patients with COVID-19-induced ITP have a wide range of clinical presentations including asymptomatic, petechiae, epistaxis, cutaneous bleeding, ICH, and cerebral haemorrhage [14, 18]. Kewan et al. reported that most of their patients with COVID-19-induced ITP had Grade 1-2 bleeding based on the modified World Health Organization (WHO) bleeding score. The clinical presentations of COVID-19-induced ITP may occur within the first week of SARS-CoV-2 infection or with delay [14].
Immunopathogenesis
The exact pathogenesis of COVID-19-induced ITP is still unclear. However, it has been suggested the SARS-CoV-2 viral antigens induce a response that cross-react with normal platelets and, in turn, cause platelet destruction or inhibit platelet release [13]. It seems that SARS-CoV-2 infection stimulates a significant immune response, given the excellent response to treatment (IVIG and corticosteroid) in patients with COVID-19-induced ITP. This viral-autoimmune response could be due to several causes such as molecular mimicry, cryptic antigen expression, or epitope spreading [3, 12].
Dysregulation of T-cells
The immunopathogenesis of ITP consists of the dysregulation in the population of T-cells which leads to the production of autoantibodies against platelets and megakaryocytes. Subsequently, cytotoxic CD8+ T-cells lyse these platelets and megakaryocytes. These two mechanisms can occur in parallel or separately. Therefore, the induction of apoptosis in platelets or megakaryocytes and their subsequent phagocytosis by macrophages and dendritic cells, increase platelet clearance and decrease platelet count [19, 20]. CD4+ T-cell imbalance also occurs in ITP in which the Th1/Th2 ratio increases in favour of the Th1 phenotype. Th1 is responsible for the response to intracellular pathogens and its overexpression can lead to autoimmune diseases. CD4 T-cells are essential for the conversion of B-cells to plasma cells; therefore, an increase in the Th1 ratio produces autoreactive B-cells and secretion of autoantibodies. Th17 is another inflammatory T-cell that is elevated in ITP, which, in turn, disrupts the Th1/Th2 ratio and increases autoantibodies [21, 22]. Another proposed mechanism in ITP is a disruption in the proportion of cytotoxic CD8+ T-cell subtypes. This induces platelet apoptosis and inhibits thrombopoiesis in megakaryocytes [20]. A summary of the dysregulation of T-cells in COVID-19-induced ITP is shown in Figure 1.
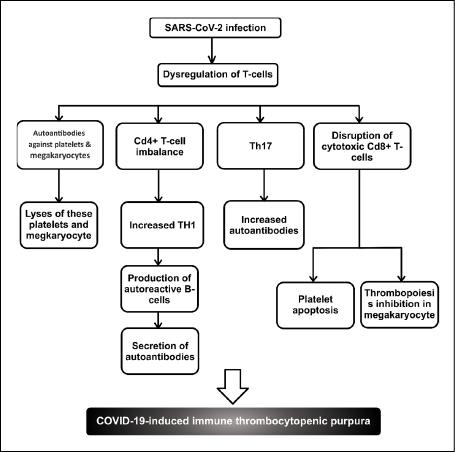
Figure 1 - Dysregulation of T-cells in COVID-19-induced immune thrombocytopenic purpura.
Molecular mimicry of platelet membrane glycoproteins
The primary cause of the autoimmune responses in ITP is unclear and may be related to genetic and environmental factors. In addition, several viral infections are associated with secondary ITP such as HIV, HCV, CMV, HSV (herpes simplex viruses), Ebola virus, and EBV. There have also been reports of thrombocytopenia due to SARS-CoV-1 infection which were generally mild. Several possible mechanisms have been proposed for the development of ITP due to infection with viruses. These include molecular mimicry of platelet membrane glycoproteins, a direct attack on megakaryocytes, induction of megakaryocyte apoptosis, and activation of inflammasomes in platelets [23, 24].
The stimulation of the membrane glycoproteins of the platelets by viruses induces CD4+ T-cell-assisted B-cell response. This induction leads to the production of antibodies (mostly IgG) against GP IIb/IIIa receptors and targets both viruses and platelets. In addition, this molecular mimicry activates CD8+ T-cells which directly causes platelet lysis [23, 24].
Direct attack on megakaryocytes and induction of megakaryocyte apoptosis
Another possible mechanism for the development of ITP, such as that seen in HIV-associated thrombocytopenia, is the direct viral attack on megakaryocytes, induction of apoptosis in megakaryocytes, and reduced platelet production. Measuring the thrombopoietin level, which is the most important regulator of thrombopoiesis, can predict and differentiate the involved mechanisms of platelet destruction or reduction due to direct viral attack on megakaryocytes [25, 26]. Scaradavou evaluated the incidence of thrombocytopenia in patients with HIV and monitored the thrombopoietin levels. He found that a lower plasma level of thrombopoietin was associated with a more normal mass of megakaryocytes in the bone marrow and platelet destruction was more involved in the mechanism of ITP. On the other hand, a higher level of thrombopoietin was associated with a lower mass of megakaryocytes and lower platelet production, and bone failure was more involved in the mechanisms of ITP [20, 26].
Hyperactivity of NLRP3-inflammasome; A key factor in COVID-19-induced immune thrombocytopenic purpura
The NOD-like receptor family pyrin domain-containing 3 (NLRP3)-inflammasome is a multi-protein complex that has a key role in detecting a variety of pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs). Activation of NLRP3-inflammasome results in an inflammatory cell death called pyroptosis as well as the production of active cytokines such as interleukin (IL)-1β and IL-18 [27, 28]. Currently, hyperactivity and dysregulation of inflammasome function, especially NLRP3-inflammasome, is considered a key mechanism in the immunopathogenesis of many inflammatory disorders such as neurodegenerative diseases, gout, and metabolic syndromes [29].
Studies on beta-coronaviruses at the onset of the COVID-19 outbreak reported that cytokine storm occurs significantly in COVID-19 patients due to the hyperactivity and dysregulation of NLRP3-inflammasome [30, 31]. Recent articles have also highlighted the potential role of NLRP3-inflammasome in COVID-19. For example, studies have investigated the role of non-coding RNAs in the regulation of NLRP3-inflammasome complex in COVID-19, the relationship between NLRP3-inflammasome and probiotics in COVID-19, the various aspects of inflammasomes as role players in COVID-19, and the potential role of the NLRP3-inflammasomes in COVID-19 [32-41]. Interestingly, a recent study has shown that specific suppression of the NLRP3-inflammasome represses the immune over-activation and diminishes COVID-19-like pathology in mice [42].
Furthermore, studies have shown the significant effects of NLRP3-inflammasome on T-cell responses. A study clearly showed that the NLRP3 gene and level of protein expression in patients with active ITP were significantly higher than in healthy controls and ITP patients in remission. Apoptosis-associated speck-like protein containing a caspase-1 recruitment domain (ASC) and caspase-1 as other major components of NLRP3-inflammasome also showed higher expression in the active ITP group compared to the control group. NLRP3 and ASC expression had a significant correlation in the active ITP group, confirming the destructive role of NLRP3-inflammasome in the occurrence of ITP. Interestingly, the plasma levels of IL-18 were higher in patients with active ITP compared to controls. On the other hand, this rate returned to normal levels in individuals that had survived. Overall, the findings of this study suggested that the activation of NLRP3-inflammasome could be a potential mechanism in the immunopathogenesis of ITP [43]. Another study reported an increase in NLRP3-, NLRC4-, and NLRP6-inflammasomes in patients with primary ITP [19]. The association of increased HMGB1 as an early detection marker with the activation of NLRP3-inflammasome activation has been demonstrated, especially in pediatric cases of ITP [44].
In a study, the polymorphisms in many inflammation-related genes associated with ITP were evaluated. In addition to SNPs of genes associated with inflammation, the NLRP3 gene polymorphism was also studied. Although no significant association between most SNPs was observed in this study, the protective role of rs10499194 was reported in ITP [45].
Another study showed that platelets regulate the activation of NLRP3-inflammasome and IL1-β production through a variety of pathways, such as platelet-derived lipid mediators, purines, nucleic acids, and a large number of platelet-derived cytokines. Therefore, any abnormality in platelet count can be associated with the dysregulation of NLRP3-inflammasome [2]. These are important during extensive cytopenia such as in COVID-19-induced ITP. One type of cell death is pyroptosis which is caused by the activation of an inflammasome complex. Although the exact role and involvement of inflammasome in patients with ITP is not fully understood, increased expression of NLRP3-inflammasome in platelets and its associated inflammatory cytokines have been observed in patients with ITP and some other autoimmune diseases. Cell destruction by HIV and many other viruses is due to the activation of the inflammasome and subsequent pyroptosis [46]. Therefore, this protein complex could be one of the possible mechanisms of ITP that develop after viral infections such as SARS-CoV-2. In addition to establishing homeostasis in innate and acquired immune systems, the platelets play an active role in controlling infections and causing inflammatory reactions. In conditions such as sepsis, malaria, or viral infection (such as HIV), platelets have been shown to produce a chain of inflammatory responses by activating the inflammasomes (predominantly NLRP3-inflammasome) [2].
The platelet-secreted IL-1β which is induced through the activation of IL-IR on endothelial cells promotes the expression of VCAM-1 (vascular cell adhesion molecule 1) in endothelial cells and, consequently, increases their permeability and plasma leakage. These are primarily a defence reaction for the migration of neutrophils and immune cells but their hyperactivity can lead to thrombocytopenia. IL-18 cytokine, also known as interferon-(IFN)-γ-inducing factor, is another important inflammatory cytokine. One of the effects of IL-18 over-activation is the increased proliferation and cytotoxic activity of CD8+ T-cells which leads to the lysis of platelets or megakaryocytes [16]. This cytokine can also change the Th1/Th2 ratio in favour of Th1. As previously stated, this imbalance is associated with increased autoreactive B-cells and secretion of autoantibodies.
Further investigations on the hyperactivity of NLRP3-inflammasome and SARS-CoV-2 infection-induced ITP are required. The previous studies highlight the importance of further studies to better understand the immunopathogenesis of SARS-CoV-2 infection and to find the missing parts of the mysterious puzzle of this disease. As a result, more effective therapeutic interventions would be eventually achieved. Given the urgent requirement for COVID-19 treatment options and considering the costs of producing new drugs, drug repurposing seems beneficial [47]. Therefore, several clinical trials have been conducted on various drugs and therapies that modulate the immune system [48, 49]. The NLRP3-inflammasome inhibitors investigated in the clinical trials as potential COVID-19 treatments include statins, sirolimus, azithromycin, cyclosporine, oridonin, quercetin, curcumin, and Tranilast [50, 51]. These drugs may have the potential to be treatment options in COVID-19-induced ITP.
A summary of the roles of platelets in COVID-19-induced ITP is shown in Figure 2. In addition, Figure 3 shows a schematic diagram of the evidence displaying the potential role of inflammasomes in the pathogenicity of ITP.
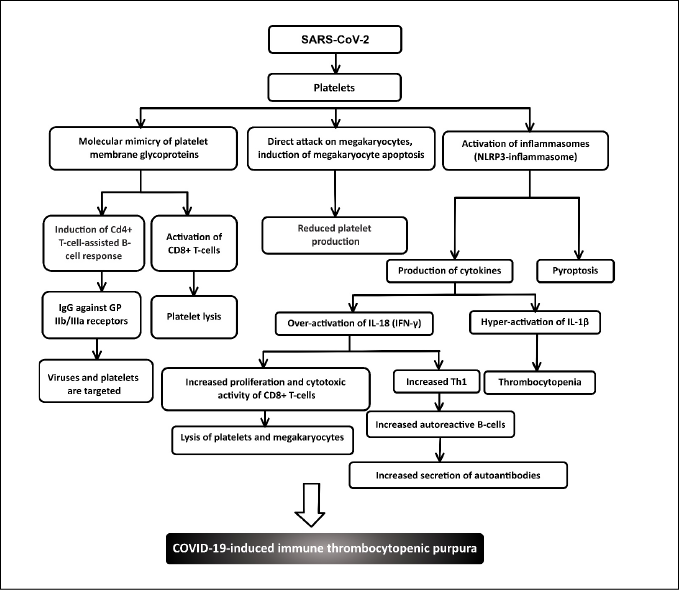
Figure 2 - Platelets in COVID-19-induced immune thrombocytopenic purpura (Ig: Immunoglobulin; GP: Glycoprotein; IL: Interleukin; INF: Interferon).
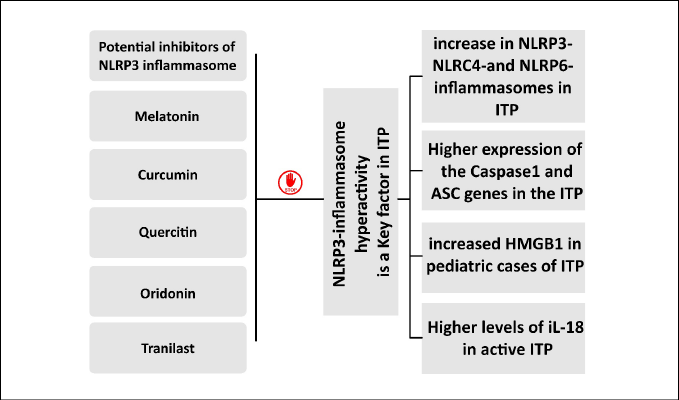
Figure 3 - Schematic diagram of the evidence displaying the potential role of inflammasomes in the pathogenicity of immune thrombocytopenic purpura (ITP) as well as the potential inhibitors of NLRP3-inflammasome.
Diagnosis
The diagnosis of ITP is established by ruling out other causes of thrombocytopenia such as thrombotic thrombocytopenic purpura (TTP), hemolytic uremic syndrome (HUS), microangiopathic hemolytic anemia (MAHA), and heparin-induced thrombocytopenia (HIT) [15, 20, 52].
Management
Patients with ITP that have severe thrombocytopenia (platelets less than 20,000) should be treated. Treatment usually involves prednisolone (1 mg/kg /day). Steroids help to prevent bleeding and reduce platelet destruction [53]. Most ITP patients respond to prednisolone treatment within two weeks and the major response is seen within the first week of treatment [53]. The management of COVID-19-induced ITP depends on the patient’s underlying condition, the extent of platelet depletion, and clinical presentations [54, 55]. Patients with ITP that do not have life-threatening hemorrhages are usually observed initially. However, following the progression of the disease, other treatment options such as corticosteroids, intravenous immunoglobulin (IVIG), anti-D Ig, and rituximab may be administered [55]. The first-line treatment of COVID-19-associated thrombocytopenia includes administration of IVIG (1g/kg) combined with systemic corticosteroids such as prednisolone or dexamethasone. To avoid the risk of secondary infection, it is best to keep the dose and duration of steroid treatment to a minimum and then initiate tapering after a maximum of two weeks. More than 70% of patients respond well to the first-line treatment but a small percentage of patients are resistant [54]. There have been reports on the effectiveness of combination therapy with dexamethasone and rituximab. The next treatment options in patients that are unresponsive to treatment include thrombopoietin receptor agonists (TPO-RAs) or splenectomy [54, 55]. TPO-RAs (such as eltrombopag) provoke platelet production by stimulating thrombopoietin (TPO) receptors on megakaryocytes [54]. Combination therapy with systemic steroids and monoclonal antibodies (such as rituximab) is recommended as the main treatment option in patients with COVID-19- associated ITP [55]. Studies have reported that patients with ITP or those receiving steroids have a risk for thrombosis [56]. In addition, there is a risk of thrombosis in patients with COVID-19. Therefore, the balance between thrombosis and bleeding must be considered when selecting the appropriate treatment option for patients with ITP. In addition, platelet transfusion should be performed alone or simultaneously with other treatments only if there is severe and life-threatening bleeding [54, 57]. A study conducted by Rampotas et al. showed that low-dose (prednisolone 20 mg) and high-dose corticosteroids had equal effectiveness. They also reported that TPO-RAs are highly effective either as the first-line or additional treatment option. Moreover, patients receiving TPO-RAs showed lower rates of thrombosis compared to patients treated with steroids or without any treatment [56]. The treatment options in COVID-19-induced ITP are shown in Table 1.

Prognosis and clinical outcome
Kewan et al. reported that excellent prognosis and clinical of outcome COVID-19-induced ITP with a response rate of 72.8% to treatment with corticosteroids and IVIG. The median time to ITP recovery in their study was four days. They also reported that none of the deaths in patients with COVID-19-induced ITP were associated with bleeding or ITP [14]. In addition, a recent systematic review and meta-analysis reported that most of the patients that had developed COVID-19-associated ITP recovered in less than one week [4]. Alonso-Beato et al. reported an overall mortality of 7% in patients with COVID-19-induced ITP. About 2.8% and 4.8% of the patients expired due to bleeding events and respiratory failure (caused by COVID-19), respectively [12].
COVID-19 vaccine-induced immune thrombocytopenic purpura
Vaccine-induced ITP has been associated with both live and inactivated vaccines (such as rubella, pneumococcus, and influenza vaccines) and has been linked to increased function of B-cells [58]. Furthermore, different types of COVID-19 vaccines have been reported to trigger de novo ITP including the Pfizer and Moderna (mRNA-based vaccines) and ChAdOx1 nCov-19 (AZD122; AstraZeneca) and Ad26.COV2. S (Janssen) (vector-based vaccines) [58, 59]. These patients had been treated with the same treatment options for COVID-19-induced ITP (such as IVIGs, glucocorticoids, or TPO-RAs) [58, 59]. In addition, there have been reports of exacerbations of ITP following COVID-19 vaccinations. Dijk et al. and Kuter et al. reported a relapse incidence of 8% and 12% in ITP patients, respectively [60, 61]. Ali et al. suggested delaying the administration of the second dose of the COVID-19 vaccine in pregnant patients with ITP flare following COVID-19 vaccination to avoid thrombocytopenia-related complications which could affect the mode of delivery [62]. The term ‘vaccine-induced immune thrombotic thrombocytopenia (VITT)’ refers to the thrombotic complications that occur following the administration of the adenovirus vector-based COVID-19 vaccines. However, the exact pathogenesis of VITT is still unclear. It has been suggested that the polyanionic constituents of the adenovirus vector-based vaccines trigger antibody production against PF4 (platelet factor 4) which, in turn, induce platelet activation. As a result, thrombocytopenia and thrombosis occur [63].
CONCLUSIONS
SARS-CoV-2 infection and ITP have very complex immunopathogenesis. The dysregulation and dysfunction of immune responses are the key factors in both conditions. However, the exact immunopathogenesis of both diseases is not yet fully understood. Various hematological events such as thrombocytopenia have been observed in SARS-CoV-2 infection. ITP is a type of thrombocytopenia that occurs due to increased platelet depletion or decreased platelet production in the bone marrow following immune responses. A greater focus on the cause of COVID-19-induced ITP not only helps to better understand the underlying immunopathogenesis and find an effective treatment for COVID-19 but also provides a new perspective on the causes of ITP, especially in other inflammatory disorders similar to COVID-19. Unregulated immune responses, especially by T lymphocytes, are the main involved mechanisms in ITP. The pathological effects of NLRP3-inflammasome hyper-activation such as hyper-inflammation through the overproduction of IL-1β and IL-18, pyroptosis, increased Th17 level, the destructive activity of IL-17, and immune dysregulation might be the causes of severe thrombocytopenia and lymphopenia in COVID-19. Further studies on the common immunopathological aspects of ITP, COVID-19, and other severe inflammatory disorders could establish a new approach to treating ITP in the future.
Acknowledgments
The authors would like to thank the Deputy of Research and Technology, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran for the support of this study.
Conflicts of interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Funding
Not applicable.
REFERENCES
[1] Sharma A, Tiwari S, Deb MK, Marty JL. Severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2): a global pandemic and treatment strategies. Int J Antimicrob Agents. 2020; 56 (2), 106054.
[2] Rolfes V, Ribeiro LS, Hawwari I, et al. Platelets Fuel the Inflammasome Activation of Innate Immune Cells. Cell Rep. 2020; 31 (6), 107615.
[3] Bhattacharjee S, Banerjee M. Immune Thrombocytopenia Secondary to COVID-19: a Systematic Review. SN Compr Clin Med. 2020; 1-11.
[4] Alharbi MG. COVID-19 associated with immune thrombocytopenia: a systematic review and meta-analysis. Expert Rev Hematol. 2022. doi: 10.1080/17474086.2022.2029699.
[5] Cines DB, Blanchette VS. Immune thrombocytopenic purpura. N Engl J Med. 2002; 346 (13), 995-1008.
[6] Rodeghiero F, Stasi R, Gernsheimer T, et al. Standardization of terminology, definitions and outcome criteria in immune thrombocytopenic purpura of adults and children: report from an international working group. Blood. 2009; 113 (11), 2386-93.
[7] Zeller B, Rajantie J, Hedlund-Treutiger I, et al. Childhood idiopathic thrombocytopenic purpura in the Nordic countries: epidemiology and predictors of chronic disease. Acta Paediatr. 2005; 94 (2), 178-84.
[8] Kupietzky A, Parnasa E, Fischer M, Kuint R, Daana M. Immune Thrombocytopenia Secondary to COVID-19 Infection. Isr Med Assoc J. 2021; 23 (6), 342-3.
[9] Audia S, Mahévas M, Samson M, Godeau B, Bonnotte B. Pathogenesis of immune thrombocytopenia. Autoimmun Rev. 2017; 16 (6), 620-32.
[10] Cines DB, Bussel JB, Liebman HA, Luning Prak ET. The ITP syndrome: pathogenic and clinical diversity. Blood. 2009; 113 (26), 6511-21.
[11] Cooper N, Bussel J. The pathogenesis of immune thrombocytopaenic purpura. Br J Haematol. 2006; 133 (4), 364-74.
[12] Alonso-Beato R, Morales-Ortega A, Fernández FJH, et al. Immune thrombocytopenia and COVID-19: Case report and review of literature. Lupus. 2021; 30 (9), 1515-21.
[13] Guirguis N, Rehman T, Shams Y, et al. SARS-CoV-2 Infection Inducing Immune Thrombocytopenic Purpura: Case Series. Ochsner J. 2021; 21 (2), 187-9.
[14] Kewan T, Gunaratne TN, Mushtaq K, Alayan D, Daw H, Haddad A. Outcomes and management of immune thrombocytopenia secondary to COVID-19: Cleveland clinic experience. Transfusion. 2021; 61 (7), 2014-8.
[15] Justiz Vaillant AA, Gupta N. ITP-Immune Thrombocytopenic Purpura. StatPearls. Treasure Island (FL): StatPearls Publishing Copyright© 2022, StatPearls Publishing LLC. 2022.
[16] Zulfiqar AA, Lorenzo-Villalba N, Hassler P, Andrès E. Immune Thrombocytopenic Purpura in a Patient with Covid-19. N Engl J Med. 2020; 382 (18), e43.
[17] Guan WJ, Ni ZY, Hu Y, et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N Engl J Med. 2020; 382 (18), 1708-20.
18. Hayashi K, Wada H, Shiraki K, Makino K. Immune Thrombocytopenia and Cerebral Hemorrhaging Associated with COVID-19. Intern Med. 2021; 60 (21), 3513.
[19] Zufferey A, Kapur R, Semple JW. Pathogenesis and Therapeutic Mechanisms in Immune Thrombocytopenia (ITP). J Clin Med. 2017; 6 (2), 16.
[20] Swinkels M, Rijkers M, Voorberg J, Vidarsson G, Leebeek FWG, Jansen AJG. Emerging Concepts in Immune Thrombocytopenia. Front Immunol. 2018; 9, 880.
[21] Ji X, Zhang L, Peng J, Hou M. T cell immune abnormalities in immune thrombocytopenia. J Hematol Oncol. 2014; 7, 72.
[22] Takahashi N, Saitoh T, Gotoh N, et al. The cytokine polymorphisms affecting Th1/Th2 increase the susceptibility to, and severity of, chronic ITP. BMC Immunol. 2017; 18 (1), 26.
[23] Franchini M, Veneri D, Lippi G. Thrombocytopenia and infections. Expert Rev Hematol. 2017; 10 (1), 99-106.
[24] Sun S, Urbanus RT, Ten Cate H, et al. Platelet Activation Mechanisms and Consequences of Immune Thrombocytopenia. Cells. 2021; 10 (12), 3386.
[25] Kühne T. Diagnosis and management of immune thrombocytopenia in childhood. Hamostaseologie. 2017; 37 (1), 36-44.
[26] Scaradavou A. HIV-related thrombocytopenia. Blood Rev. 2002; 16 (1), 73-6.
[27] Kelley N, Jeltema D, Duan Y, He Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int J Mol Sci. 2019; 20 (13), 3328.
[28] de Zoete MR, Palm NW, Zhu S, Flavell RA. Inflammasomes. Cold Spring Harb Perspect Biol. 2014; 6 (12), a016287.
[29] Yang Y, Wang H, Kouadir M, Song H, Shi F. Recent advances in the mechanisms of NLRP3 inflammasome activation and its inhibitors. Cell Death Dis. 2019; 10 (2), 128.
[30] Bertocchi I, Foglietta F, Collotta D, et al. The hidden role of NLRP3 inflammasome in obesity-related COVID-19 exacerbations: Lessons for drug repurposing. Br J Pharmacol. 2020; 177 (21), 4921-30.
[31] van den Berg DF, Te Velde AA. Severe COVID-19: NLRP3 Inflammasome Dysregulated. Front Immunol. 2020; 11, 1580.
[32] Gedefaw L, Ullah S, Lee TMH, Yip SP, Huang CL. Targeting Inflammasome Activation in COVID-19: Delivery of RNA Interference-Based Therapeutic Molecules. Biomedicines. 2021; 9 (12), 1823.
[33] Houshmandfar S, Saeedi-Boroujeni A, Rashno M, Khodadadi A, Mahmoudian-Sani MR. miRNA-223 as a regulator of inflammation and NLRP3 inflammasome, the main fragments in the puzzle of immunopathogenesis of different inflammatory diseases and COVID-19. Naunyn Schmiedebergs Arch Pharmacol. 2021; 394 (11), 2187-95.
[34] Kasti AN, Synodinou KD, Pyrousis IA, Nikolaki MD, Triantafyllou KD. Probiotics Regulating Inflammation via NLRP3 Inflammasome Modulation: A Potential Therapeutic Approach for COVID-19. Microorganisms. 2021; 9 (11), 2376.
[35] Amin S, Aktar S, Rahman MM, Chowdhury MMH. NLRP3 inflammasome activation in COVID-19: an interlink between risk factors and disease severity. Microbes Infect. 2022; 24 (1), 104913.
[36] Bazrafkan M, Hosseini E, Nazari M, Amorim CA, Sadeghi MR. NLRP3 inflammasome: A joint, potential therapeutic target in management of COVID-19 and fertility problems. J Reprod Immunol. 2021; 148, 103427.
[37] Corpetti C, Del Re A, Seguella L, et al. Cannabidiol inhibits SARS-CoV-2 spike (S) protein-induced cytotoxicity and inflammation through a PPARγ-dependent TLR4/NLRP3/Caspase-1 signaling suppression in Caco-2 cell line. Phytother Res. 2021; 35 (12), 6893-903.
[38] Manayi A, Nabavi SM, Khayatkashani M, Habtemariam S, Khayat Kashani HR. Arglabin could target inflammasome-induced ARDS and cytokine storm associated with COVID-19. Mol Biol Rep. 2021; 48 (12), 8221-5.
[39] Shohan M, Nashibi R, Mahmoudian-Sani MR, et al. The therapeutic efficacy of quercetin in combination with antiviral drugs in hospitalized COVID-19 patients: A randomized controlled trial. Eur J Pharmacol. 2022; 914, 174615.
[40] Vora SM, Lieberman J, Wu H. Inflammasome activation at the crux of severe COVID-19. Nat Rev Immunol. 2021; 21 (11), 694-703.
[41] Yalcinkaya M, Liu W, Islam MN, et al. Modulation of the NLRP3 inflammasome by Sars-CoV-2 Envelope protein. Sci Rep. 2021; 11 (1), 24432.
[42] Zeng J, Xie X, Feng XL, et al. Specific inhibition of the NLRP3 inflammasome suppresses immune overactivation and alleviates COVID-19 like pathology in mice. E Bio Medicine. 2022; 75, 103803.
[43] Qiao J, Liu Y, Li X, et al. Elevated expression of NLRP3 in patients with immune thrombocytopenia. Immunol Res. 2016; 64 (2), 431-7.
[44] Wang R, Cao Q, Bai ST, Wang L, Sheng GY. Potential role and mechanism for high mobility group box1 in childhood chronic immune thrombocytopenia. Eur Rev Med Pharmacol Sci. 2019; 23 (24), 10931-41.
[45] Li J, Ma S, Shao L, et al. Inflammation-Related Gene Polymorphisms Associated With Primary Immune Thrombocytopenia. Front Immunol. 2017; 8, 744.
[46] Doitsh G, Galloway NL, Geng X, et al. Cell death by pyroptosis drives CD4 T-cell depletion in HIV-1 infection. Nature. 2014; 505 (7484), 509-14.
[47] Mule S, Singh A, Greish K, Sahebkar A, Kesharwani P, Shukla R. Drug repurposing strategies and key challenges for COVID-19 management. J Drug Target. 2021; 1-17.
[48] Alunno A, Najm A, Mariette X, et al. Immunomodulatory therapies for SARS-CoV-2 infection: a systematic literature review to inform EULAR points to consider. Ann Rheum Dis. 2021; 80 (6), 803-15.
[49] Burrage DR, Koushesh S, Sofat N. Immunomodulatory Drugs in the Management of SARS-CoV-2. Front Immunol. 2020; 11, 1844.
[50] Parsamanesh N, Moossavi M, Bahrami A, Fereidouni M, Barreto G, Sahebkar A. NLRP3 inflammasome as a treatment target in atherosclerosis: A focus on statin therapy. Int Immunopharmacol. 2019; 73, 146-55.
[51] Saeedi-Boroujeni A, Mahmoudian-Sani MR. Anti-inflammatory potential of Quercetin in COVID-19 treatment. J Inflamm (Lond). 2021; 18 (1), 3.
[52] Lévesque V, Millaire É, Corsilli D, Rioux-Massé B, Carrier FM. Severe immune thrombocytopenic purpura in critical COVID-19. Int J Hematol. 2020; 112 (5), 746-50.
[53] Stasi R, Provan D. Management of immune thrombocytopenic purpura in adults. Mayo Clin Proc. 2004; 79 (4), 504-22.
[54] Pavord S, Thachil J, Hunt BJ, et al. Practical guidance for the management of adults with immune thrombocytopenia during the COVID-19 pandemic. Br J Haematol. 2020; 189 (6), 1038-43.
[55] Ahmed N, Asreb A, Chofor R, Melese A. Treatment of Severe Immune Thrombocytopenic Purpura Associated with COVID-19. Am J Case Rep. 2021; 22, e932557.
[56] Rampotas A, Watson E, Burton K, Hill QA, Pavord S. A real-world study of immune thrombocytopenia management during the COVID-19 pandemic in the UK. Br J Haematol. 2022; 196 (2), 351-5.
[57] Kewan T, Almhana F, Schwartzman L, Daw H, Haddad A. COVID-19 patient with immune thrombocytopenic purpura. Int J Lab Hematol. 2020; 42 (6), e260-e2.
[58] Paulsen FO, Schaefers C, Langer F, et al. Immune thrombocytopenic purpura after vaccination with COVID-19 vaccine (ChAdOx1 nCov-19). Blood. 2021; 138 (11), 996-9.
[59] Lee EJ, Cines DB, Gernsheimer T, et al. Thrombocytopenia following Pfizer and Moderna SARS-CoV-2 vaccination. Am J Hematol. 2021; 96 (5), 534-7.
[60] Dijk W, Schutgens REG. Relapse of immune thrombocytopenia after COVID-19 vaccination. Eur J Haematol. 2022; 108 (1), 84-5.
[61] Kuter DJ. Exacerbation of immune thrombocytopenia following COVID-19 vaccination. Br J Haematol. 2021; 195 (3), 365-70.
[62] Ali E, Al-Maharmeh Q, Rozi WM, Habib MB, Yassin M. Immune thrombocytopenia purpura flare post COVID-19 vaccine. Ann Med Surg (Lond). 2021; 103164.
[63] Thilagar B, Beidoun M, Rhoades R, Kaatz S. COVID-19 and thrombosis: searching for evidence. Hematology Am Soc Hematol Educ Program. 2021; 2021 (1), 621-7.