Le Infezioni in Medicina, n. 2, 181-190, 2021
REVIEWS
Molecular and serological prevalence of Coronavirus in Chiropterans: A systematic review with meta-analysis
D. Katterine Bonilla-Aldana1,2, Doménica Acevedo-López3,4,
Brayan Stiven Aristizábal-Carmona3,4, Fabio Andrés Díaz-García3,4, Cristian Camilo Sarmiento-Cano3,4, Samuel Gutiérrez-Soleibe3,4, María Del Mar España-Cerquera3,4, Diego Mauricio Obando-Cardona3,4, Laura Paulina Castrillón-Correa3,4, Juliana Castro-Henao3,4, Juan Esteban Suárez-Muñoz3,4, Sara Cristina Serna-Suárez3,4, Mateo Alejandro Mora-Hernández3,4, Valeria Álvarez-Amaya3,4, Alfonso J. Rodriguez-Morales2,5,6,7, Samuel Pecho-Silva7,8, Alberto Paniz-Mondolfi9,10, Salim Mattar11
1Semillero de Investigación en Zoonosis (SIZOO), Grupo de Investigación BIOECOS, Fundación Universitaria Autónoma de las Américas, Sede Pereira, Pereira, Risaralda, Colombia;
2Committee on Tropical Medicine, Zoonoses and Travel Medicine, Asociación Colombiana de Infectología, Bogotá, DC, Colombia;
3Asociación Científica de Estudiantes de Medicina de la Fundación Universitaria Autónoma de las Américas (ASCEMAP), Pereira, Risaralda, Colombia;
4Faculty of Medicine, Fundación Universitaria Autónoma de las Américas, Pereira, Risaralda, Colombia;
5Grupo de Investigación Biomedicina, Faculty of Medicine, Fundación Universitaria Autónoma de las Américas, Pereira, Risaralda, Colombia;
6School of Medicine, Universidad Privada Franz Tamayo, Cochabamba, Bolivia.
7Universidad Científica del Sur, Lima, Peru;
8Hospital Nacional Edgardo Rebagliati Martins, Lima, Perú;
9Department of Pathology, Molecular and Cell-Based Medicine, Laboratory of Microbiology, Icahn School
of Medicine at Mount Sinai, New York, New York, USA;
10Instituto de Investigaciones Biomédicas IDB/Incubadora Venezolana de la Ciencia, Barquisimeto, Venezuela;
11Instituto de Investigaciones Biologicas del Tropico, Universidad de Cordoba, Monteria, Colombia
Corresponding author
Alfonso J. Rodriguez-Morales
E-mail: alfonso.rodriguez@uam.edu.co
arodriguezmo@cientifica.edu.pe
SummaRY
In recent years, and now especially with the arrival of the Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2), there has been increased interest in understanding the role of bats in the dynamics of transmission and origin of this pandemic agent. To date, no systematic reviews have been published on this topic. This systematic review aimed to summarize and highlight the frequency of bat infections reported in currently available observational studies for coronavirus. The purpose of this study was also to examine the differences between the pool prevalence by technique and country. We performed a systematic literature review with meta-analysis, using three databases to assess coronavirus (CoV) infection in bats and its diagnosis by serological and molecular tests. We carried out random-effects model meta-analysis to calculate the pooled prevalence and 95% confidence interval (95% CI). In all, 824 articles were retrieved (1960-2021). After screening by abstract/title, 43 articles were selected for full-text assessment. Of these, 33 were finally included for qualitative and quantitative analyses. From the total of studies, the pool prevalence by RT-PCR (n=14,295 bats) for CoV was 9.8% (95% CI 8.7-10.9%); Italy reported the highest pooled prevalence (44.9%, 95% CI 31.6-58.1%), followed by the Philippines (29.6%). Regarding the ELISA, the pool prevalence for coronavirus from 15 studies, including 359 bats, was 30.2% (95% CI 14.7-45.6%). The results for coronaviruses with the MIF were significantly lower, 2.6% (95% CI 1.5-3.7%). A considerable proportion of infected bats tested positive, particularly by molecular tests. This essential condition highlights the relevance of bats and the need for future studies to detail their role as potential reservoirs of SARS-CoV-2. In this meta-analysis, bats were positive in almost 10% by RT-PCR, suggesting their relevance and the need to understand their potential participation in maintaining wild zoonotic transmission.
Keywords: Bats; coronavirus; SARS-CoV-2; systematic review; meta-analysis.
INTRODUCTION
Bats are recognized for being a potential reservoir of various pathogens due to their high diversity and multiple cell types and receptors [1]. After humans and rodents, bats are the most abundant of mammals. They are essential for maintaining the environment, and their guano is used as fertilizer and for diverse manufacturing. Depending on the species, their diet is based on insects, small mammals, fish, blood, fruits and pollen. Many viruses can be found in bats, but not all of these viruses can be transmitted to humans. The most common viruses leading to zoonosis are Nipah, Hendra, Ebola, and rabies, although other hosts are primary and more relevant in transmission, e.g. dogs for rabies. Now Coronaviruses, due to the Coronavirus Disease 2019 (COVID-19) pandemic, are under consideration as needed for research on the links with them [2-5].
The Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2), the etiological agent of COVID-19, is a relatively recent virus that first appeared in the Chinese seafood and animal markets in Wuhan. Since then, it has caused more than 2.3 million deaths worldwide [6-8]. This new virus was identified from bronchoalveolar lavage samples from patients with pneumonia in China [7, 9-11]. It was found to be part of the coronavirus family, more specifically the betacoronavirus genus, related to SARS-CoV that caused the Severe Acute Respiratory Syndrome (SARS) in Guangdong in 2002 and caused 774 deaths in 37 different countries and Middle East Respiratory Syndrome (MERS-CoV) that appeared in Saudi Arabia in 2012 and caused 858 deaths [12, 13].
Coronaviruses infect a wide range of animals such as camels, rabbits, and mainly–apparently- several different species of bats [14]. Current scientific evidence proposes that SARS-CoV-2 has a zoonotic origin, and everything suggests to its natural reservoir being possibly bats [15]. It seems that its incredible infectious ability is because it has a Receptor-Binding Domain (RBD) that binds with greater affinity to Angiotensin-Converting Enzyme 2 (ACE2) than SARS-CoV; this may shed light on the animal-to-human transmission potential of SARS-CoV-2 [16].
In humans, this virus can present with a wide variety of presentations, from mild symptoms to severe pneumonia. It also causes an increase in morbidity and mortality rates and causes considerable expenses in the health system [17, 18]. Due to deforestation, economic development, and climate change, human interactions with these zoonotic hosts are expected to increase. That is why knowledge of the natural and intermediate reservoirs of the virus could help us to control and prevent related infectious diseases in humans, avoiding possible outbreaks [19].
It is essential to mention that still, as of April 2021, there is a lack of data in the prevalence of SARS-CoV-2 in animals. Some studies have focused on the phylogenetic analyses of SARS-CoV-2 regarding animals, such as bats and pangolins [20-22]. Some case reports and small series of natural infection of SARS-CoV-2, especially in cats, have also been reported. A systematic review of SARS-CoV-2 infection in animals and, in general, coronaviruses among bats is highly expected soon [23,24]. The aim of this systematic review was to summarize and highlight the frequency of bats infections reported on currently available observational studies for coronavirus. Also, this study aimed to examine the differences between the pool prevalence by technique and country.
MATERIAL AND METHODS
Protocol
This protocol follows the recommendations established by the PRISMA statement [25].
Eligibility criteria
We included published peer-reviewed articles that reported infection in animals with serological or molecular confirmation of Coronaviruses. We considered serological tests as Enzyme-Linked Immunosorbent Assay (ELISA) and the macrophage Migration Inhibitory Factor test (MIF), and for molecular-based testing, the Reverse Transcription Polymerase Chain Reaction (RT-PCR). The article language limit was not set, and we included publications from January 1st, 1990, until the date the search was finalized and completed (February 1st, 2021). Review articles, opinion articles, and letters not presenting original data were excluded from the study, as were studies reporting on cases with incomplete information.
Information sources and search strategy
We conducted a systematic review using Medline/PubMed, Scopus, and Web of Sciences. The search terms used were as follows: “coronavirus,” “bats”, “Chiroptera”. The searches were concluded by February 1st, 2021, and four different researchers independently evaluated the search results.
Study selection
Results of the initial search strategy were first screened by title and abstract. The full texts of relevant articles were examined for inclusion and exclusion criteria. When an article reported the same information from the same patient, information from both reports was combined to obtain complementary data, counting only as a single case. Observational studies reported the frequency of animals infected due to any coronavirus included for quantitative synthesis (meta-analysis).
Data collection process and data items
Data extraction forms, including information on the type of publication, publishing institution, country, year and date of publication, and number of infected animals assessed by serological or molecular tests, were filled independently by four investigators. A fifth researcher checked the article list and data extractions to ensure no duplicate articles or duplicate information of the same study and resolved discrepancies about study inclusion.
Assessment of methodological quality and risk of bias
For quality assessment, we used the IHE’s Quality Appraisal of Case Series Studies Checklist and the critical appraisal tool to assess the quality of cross-sectional studies (AXIS) [26, 27]. Publication bias was assessed using a funnel plot. A random-effects model was used to calculate the pooled prevalence and 95% CI, given variable degrees of data heterogeneity.
Statistical approach
Unit discordance for variables was resolved by converting all units to a standard measurement for that variable. Percentages and means ± standard deviation (SDs) were calculated to describe the distributions of categorical and continuous variables, respectively. Since individual patient information was not available for all patients, we report weighted means and SDs. The baseline data were analyzed using the Stata version 14.0, licensed for Universidad Tecnológica de Pereira.
The meta-analyses were performed using Stata® and the software Open Meta[Analyst] and Comprehensive Meta-Analysis ve.3.3® licensed for Universidad Tecnológica de Pereira [28]. Pooled prevalences and their 95% confidence intervals (95% CIs) were used to summarize the weighted effect size for each study grouping variable using the binary random-effects model (the weighting took into consideration the sample sizes of the individual studies), except for median age, where a continuous random-effect model was applied (DerSimonian-Laird procedure) [29, 30]. A random-effects meta-analysis model assumes that the effects estimated in different studies are not identical but follow some distribution. For random-effects analyses, the pooled estimate and 95%CIs refer to the centre of the pooled prevalence distribution but do not describe the width of the distribution. Often the pooled estimate and its 95%CI are quoted in isolation as an alternative estimate of the quantity evaluated in a fixed-effect meta-analysis, which is inappropriate. The 95%CI from a random-effects meta-analysis describes uncertainty in the mean of systematically different prevalence in the different studies.
Measures of heterogeneity, including Cochran’s Q statistic, the I2 index, and the tau-squared test, were estimated and reported. We performed subgroup analyses using techniques, animals, countries, and meta-analyses for each of the variables of interest. Publication bias was assessed using a funnel plot. A random-effects model was used to calculate the pooled prevalence and 95% CI, given variable degrees of data heterogeneity.
RESULTS
Study selection and characteristics
A total of 824 articles were retrieved using the search strategy. After screening by abstract and title, 43 articles were finally selected for full-text assessment. Of these, ten were excluded due to lack of information on molecular diagnosis, and 33 were finally included for the final qualitative and quantitative meta-analysis (Figure 1). The main characteristics of the included studies are shown in Tables 1, 2 and 3.
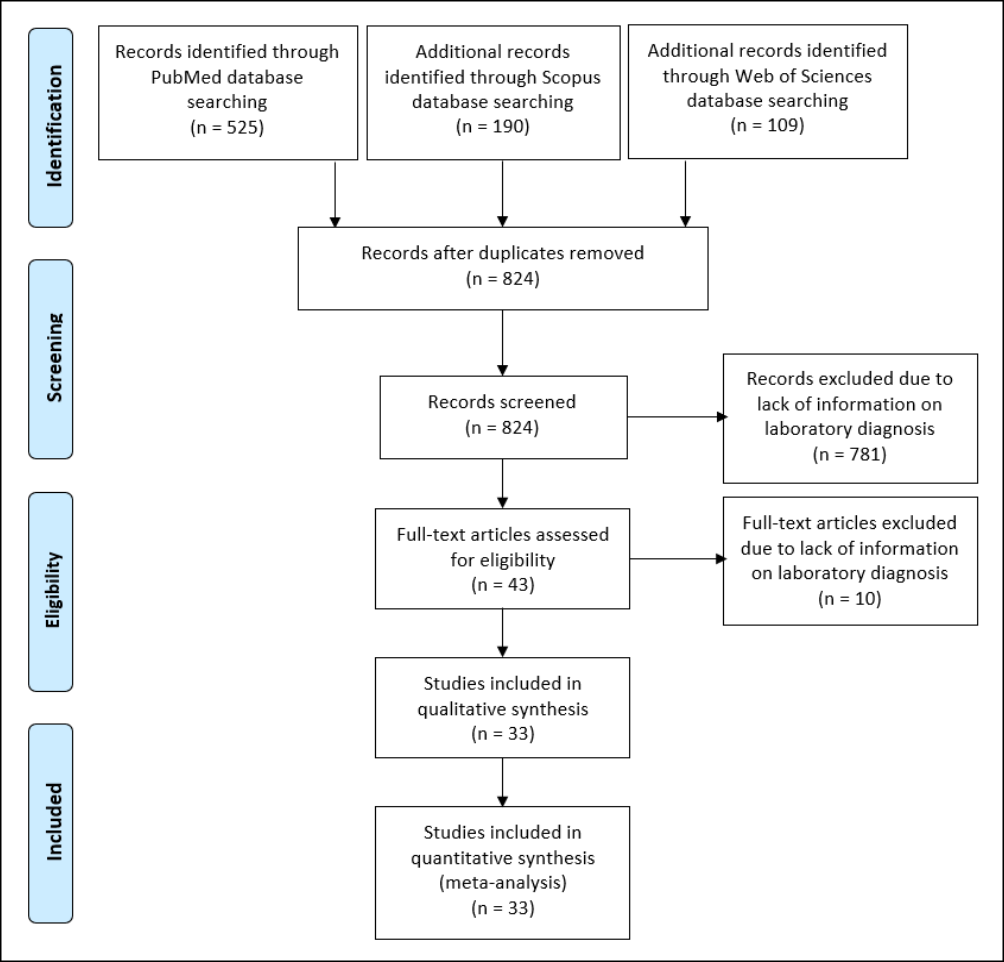
Figure 1 - Study selection and characteristics.
Table 1 - Characteristics of the studies evaluating with MIF, Coronavirus in Chiroptera.
|
Number/ code |
Title Study |
Year- Publication |
Years- Study |
State or City or Place |
Country |
Study Type |
Family |
Scientific Name |
N |
n(+) |
% |
|
RSM-02 |
Viral Diversity of Microbats within the South West Botanical Province of Western Australia |
2019 |
2016-2018 |
North of Australia |
Australia |
Prevalence |
Vespertilio-nidae |
Chalinolobus gouldii |
66 |
2 |
3.03 |
|
Chalinolobus morio |
59 |
4 |
6.78 |
||||||||
|
Falsistrellus mackenziei |
7 |
0 |
0.00 |
||||||||
|
Nyctophilus geoffroyi |
26 |
0 |
0.00 |
||||||||
|
Nyctophilus gouldi |
67 |
7 |
10.45 |
||||||||
|
Nyctophilus major |
5 |
2 |
40.00 |
||||||||
|
Scotorepens balstoni |
2 |
0 |
0.00 |
||||||||
|
Vespadelus baverstocki |
1 |
0 |
0.00 |
||||||||
|
Vespadelus regulus |
150 |
9 |
6.00 |
||||||||
|
Molossidae |
Austronomus australis |
11 |
1 |
9.09 |
|||||||
|
Ozimops sp |
1 |
0 |
0.00 |
||||||||
|
Vespertilio-nidae |
Chalinolobus gouldii |
68 |
2 |
2.94 |
|||||||
|
Chalinolobus morio |
60 |
1 |
1.67 |
||||||||
|
Falsistrellus mackenziei |
7 |
0 |
0.00 |
||||||||
|
Nyctophilus geoffroyi |
27 |
0 |
0.00 |
||||||||
|
Nyctophilus gouldi |
70 |
0 |
0.00 |
||||||||
|
Nyctophilus major |
4 |
0 |
0.00 |
||||||||
|
Scotorepens balstoni |
2 |
0 |
0.00 |
||||||||
|
Vespadelus baverstocki |
1 |
0 |
0.00 |
||||||||
|
Vespadelus regulus |
155 |
4 |
2.58 |
||||||||
|
Molossidae |
Austronomus australis |
11 |
0 |
0.00 |
|||||||
|
Ozimops sp |
1 |
0 |
0.00 |
Table 2 - Characteristics of the studies evaluating with ELISA, Coronavirus in Chiroptera.
|
Code |
Title Study |
Year |
State or City or Place |
Country |
Family |
Scientific Name |
N |
n(+) |
% |
|
|
Publication |
Study |
|||||||||
|
RSM-11 |
Bats Are Natural Reservoirs of SARS-Like Coronaviruses. |
2005 |
2004-2005 |
Nanning, Guangxi |
China |
Pteropodidae |
Rousettus leschenaulti |
84 |
1 |
1.19 |
|
RSM-11 |
2005 |
2004-2005 |
Maoming, Guangdong |
China |
Pteropodidae |
Rousettus leschenaulti |
42 |
0 |
0.00 |
|
|
RSM-11 |
2005 |
2004-2005 |
Maoming, Guangdong |
China |
Pteropodidae |
Cynopterus sphinx |
17 |
0 |
0.00 |
|
|
RSM-11 |
2005 |
2004-2005 |
Yichang, Hubei |
China |
Rhinolophidae |
Rhinolophus ferrumequinum |
4 |
0 |
0.00 |
|
|
RSM-11 |
2005 |
2004-2005 |
Yichang, Hubei |
China |
Rhinolophidae |
Rhinolophus macrotis |
7 |
5 |
71.43 |
|
|
RSM-11 |
2005 |
2004-2005 |
Yichang, Hubei |
China |
Vespertilionidae |
Nyctalus plancyi |
1 |
0 |
0.00 |
|
|
RSM-11 |
2005 |
2004-2005 |
Yichang, Hubei |
China |
Vespertilionidae |
Miniopterus schreibersi |
1 |
0 |
0.00 |
|
|
RSM-11 |
2005 |
2004-2005 |
Yichang, Hubei |
China |
Vespertilionidae |
Myotis altarium |
1 |
0 |
0.00 |
|
|
RSM-11 |
2005 |
2004-2005 |
Nanning, Guangxi |
China |
Pteropodidae |
Rousettus leschenaulti |
58 |
1 |
1.72 |
|
|
RSM-11 |
2005 |
2004-2005 |
Nanning, Guangxi |
China |
Rhinolophidae |
Rhinolophus pearsoni |
46 |
13 |
28.26 |
|
|
RSM-11 |
2005 |
2004-2005 |
Nanning, Guangxi |
China |
Rhinolophidae |
Rhinolophus pussilus |
6 |
2 |
33.33 |
|
|
RSM-25 |
Coronavirus Infection and Diversity in Bats in the Australasian Region |
2016 |
2006-2009 |
Multiple country |
Vespertilionidae |
Scotorepens spp. |
24 |
24 |
100.00 |
|
|
RSM-25 |
2016 |
2006-2009 |
Multiple country |
Vespertilionidae |
Scotophilus spp. |
4 |
4 |
100.00 |
||
|
RSM-25 |
2016 |
2006-2009 |
Multiple country |
Vespertilionidae |
Miniopterus australis |
1 |
1 |
100.00 |
||
|
RSM-25 |
2016 |
2006-2009 |
|
Multiple country |
Megadermatidae |
Macroderma gigas |
63 |
1 |
1.59 |
|
Table 3 - Meta-analysis outcomes (random-effects model)*.
|
Technique |
Number of Studies |
Pool Prevalence (%) |
95%CI |
n |
Q† |
I2 ‡ |
t2 § |
p |
|
15 |
30.2 |
14.7-45.6 |
359 |
1183.54 |
98.82 |
0.072 |
<0.001 |
|
|
MIF |
22 |
2.6 |
1.5-3.7 |
801 |
20.554 |
0.001 |
0.001 |
<0.001 |
|
RT-PCR |
33 |
9.8 |
8.7-10.9 |
14,295 |
1755.99 |
88.49 |
0.003 |
<0.001 |
|
Countries |
||||||||
|
Italy |
4 |
44.9 |
31.6-58.1 |
50 |
– |
– |
– |
– |
|
Philippines |
1 |
29.6 |
22.9-36.3 |
179 |
– |
– |
– |
– |
|
China |
21 |
28.1 |
23.0-33.1 |
4317 |
– |
– |
– |
– |
|
United Kingdom |
8 |
23.5 |
7.9-39.1 |
112 |
– |
– |
– |
– |
* 95% CI = 95% confidence interval. † Cochran’s Q statistic for heterogeneity. ‡ I2 index for the degree of heterogeneity. § Tau-squared measure of heterogeneity. ELISA, enzyme-linked immunosorbent assay; MIF, macrophage migration inhibitory factor test; RT-PCR, reverse transcription-polymerase chain reaction.
Our review included 33 studies that were published between January 1st, 2002, and February 1st, 2021, most of them from China (34%), France (6%), and Italy (6%), among others (Tables 1-3), including a total of 14,295 bats assessed by RT-PCR, 801 by MIF, and 359 by ELISA. All the studies were cross-sectional (Tables 1-3).
Individual study characteristics
The number of included animals per study was 23 for RT-PCR, 11 for MIF, and 7 for ELISA, with positive rates ranging from 0 to 100%.
Serological findings
The results for coronaviruses with the MIF were significantly lower, 2.6% (95%CI 1.5-3.7%) (Tables 1 and 3, and Figure 2). Regarding the ELISA, the pool prevalence for coronavirus from 15 studies, including 359 bats, was 30.2% (95%CI 14.7-45.6%) (Tables 2 and 3 and Figure 2).
Molecular findings
Regarding the RT-PCR, the pool prevalence for coronaviruses derived from 33 studies, including 14,295 bats, was 9.8% (95%CI 8.7-10.9%) (Figure 2).
Comparing the findings by countries, Italy reported the highest pooled prevalence (44.9%, 95%CI 31.6-58.1%), followed by the Philippines (29.6%), China (28.1%) and United Kingdom (23.5%), among others (Table 3).
DISCUSSION
Over the last decades, the interest in studying the infectious agents of bats has increased [31, 32]. Bats may harbour parasites, bacteria, fungi and viruses [33-35]. In recent years, the interest in the role of Chiroptera has been intense, especially after the SARS-CoV epidemic in 2002 in China and the MERS-CoV epidemic in 2012 in Saudi Arabia and other Middle East countries [36-42]. Now, SARS-CoV-2 affects more than 140 million people globally, with more than 3 million people dead as a result (April 17th, 2021). Despite that, there is a lack of systematic reviews about coronavirus in Chiropterans [23, 43]. Perhaps, in this systematic review, few studies (only 33) were included for the final analysis since a large number of them were classified as containing lack information on laboratory diagnosis. This point could be a reminder to the publishers of medical and scientific journals to be careful in demanding more complete and rigorous information from the authors of manuscripts in this regard [44].
In the current systematic review, we found almost one third of the assessed bats seropositive to coronaviruses, with nearly 10% having molecular detection of at least one coronavirus. Although even the detection should be carefully interpreted, as this does not necessarily mean that it is an infectious source or a reservoir, it does call on the need for further studies to confirm the fundamental role of these flying mammals in the natural cycles of coronavirus diseases such as SARS, MERS and the recently emerged SARS-CoV-2 [4, 8, 45]. Several ongoing studies are trying to describe the real risk posed by the zoonotic origin of the pandemic SARS-CoV-2 causing the COVID-19 [31, 46-52].
Although SARS-CoV-2 originated in China, as has been widely described, few efforts to study this coronavirus in bats in other countries and continents have been employed to date after one year of the pandemic. Nevertheless, some studies, among those included in the current review, have been made in other countries, including some in Asia, such as the Philippines, or Europe, as is Italy and United Kingdom. Bats are mammals widely distributed globally, and the hypotheses that all of them may harbour coronaviruses are entirely plausible [31, 32]. Recently, transmission from humans to other animals has been detected, especially in different feline animals (cats, lions, tigers), but can bats infect these animals in the wild ecotone settings [12, 13, 44]? That is a question remaining to be answered. Some experts are concerned that SARS-CoV-2/COVID-19 become endemic for multiple reasons, including the potential animal reservoirs, as well as wild species, like Chiroptera [53].
Indeed, more studies are required, especially from some areas of the world, as is the case of Africa and Latin America, where although significantly affected by millions of cases, still have not developed a significant number of studies in the One Health interphases of human, animal and environmental health in the context of coronaviruses such as the SARS-CoV-2 [54-56]. In countries such as Brazil and Mexico, with a vast jungle threated by often anthropogenic wildfires, experts are concerned about the SARS-CoV-2, and other new coronaviruses may emerge from wild animals [57-62]. There is a consensus that the question is not “if” novel zoonotic viruses will rise but “when” [62]. More research in these areas is crucial for future epidemics or even pandemic by emerging coronaviruses.
More studies, going deep into the wild cycles, bats and other different animals involved, the ecoepidemiological conditions and the areas with high risk for human interaction, are essential among all these settings. Coronaviruses are present in bats and require more assessments to define their real role in human and animal diseases. As limitation, the number of studies specifying the specific CoVs investigated is limited and techniques not uniform at all. Then, more studies are needed [63].
Author contributions
AJRM and DKBA formulated the research questions, designed the study, developed the preliminary search strategy, and drafted the manuscript. DAL, BSAC, FADG, CCSC, SGS, MDMEC, DMOC, LPCC, JCH, JESM, SCSS, MAMH, VAM refined the search strategy by conducting iterative database queries and incorporating new search terms. DAL, BSAC, FADG, CCSC, SGS, MDMEC, DMOC, LPCC, JCH, JESM, SCSS, MAMH, VAM, AJRM, and DKBA searched and collected the articles. APM, SM, SPS, AJRM, and DKBA conducted the quality assessment. All authors critically reviewed the manuscript for relevant intellectual content. All authors have read and approved the final version of the manuscript.
Conflict of interest
All authors report no potential conflicts.
Funding source
None.
Ethical approval
Approval was not required.
Acknowledgements
Authors would like to thanks the support of the Scientific Association of Medical Students of the Fundación Autónoma de las Américas (ASCEMAP), as well as to the Incubator in Zoonosis (Semillero en Zoonosis, SIZOO).
REFERENCES
[1] Woo PCY, and Lau SKP. Viruses and Bats. Viruses. 2019; 11 (10), 884.
[2] Cimerman S, Chebabo A, Cunha CAD, and Rodriguez-Morales AJ. Deep impact of Covid-19 in the healthcare of Latin America: The Case of Brazil. Braz J Infect Dis. 2020; 24 (2), 93-5.
[3] Rodriguez-Morales AJ, Katterine Bonilla-Aldana D, Tiwari R, Sah R, Rabaan AA, and Dhama K. Covid-19, an emerging coronavirus infection: current scenario and recent developments - an overview. J. Pure Appl. Microbiol. 2020; 14 (1), 5-12.
[4] Calisher CH, Childs JE, Field HE, Holmes KV, and Schountz T. Bats: Important reservoir hosts of emerging viruses. Clin Microbiol Rev. 2006; 19 (3), 531-45.
[5] Ahmad T, Khan M, Haroon, et al. Covid-19: zoonotic aspects. Travel Med Infect Dis. 2020; 36, 101607.
[6] Organization WH. “Who Coronavirus Disease (Covid-19) Dashboard.” World Health Organization, https://covid19.who.int/.
[7] Zhu N, Zhang D, Wang W, et al. A novel coronavirus from patients with pneumonia in China, 2019. N Engl J Med. 2020; 382 (8), 727-33.
[8] Dhama K, Khan S, Tiwari R, et al. Coronavirus Disease 2019-Covid-19. Clin Microbiol Rev. 2020; 33 (4), e00028-20.
[9] Halfmann PJ, Hatta M, Chiba S, et al. Transmission of Sars-Cov-2 in domestic cats. N Engl J Med. 2020; 383 (6), 592-4.
[10] Holshue ML, DeBolt C, Lindquist S, et al. First case of 2019 Novel Coronavirus in the United States. N Engl J Med. 2020; 382 (10), 929-36.
[11] Zou L, Ruan F, Huang M, et al. Sars-Cov-2 viral load in upper respiratory specimens of infected patients. N Engl J Med. 2020; 382 (12), 1177-9.
[12] Lu R, Zhao X, Li J, et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: implications for virus origins and receptor binding. The Lancet. 2020; 395 (10224), 565-74.
[13] Malik YS, Kumar N, Sircar S, et al. Coronavirus Disease Pandemic (Covid-19): challenges and a global perspective. Pathogens. 2020; 9 (7), 519.
[14] Lau SK, Poon RW, Wong BH, et al. Coexistence of different genotypes in the same bat and serological characterization of rousettus bat coronavirus Hku9 belonging to a novel betacoronavirus Subgroup. J Virol. 2010; 84 (21), 11385-94.
[15] Andersen KG, Rambaut A, Lipkin WI, Holmes EC, and Garry RF. The proximal origin of Sars-Cov-2. Nat Med. 2020; 26 (4), 450-2.
[16] Shang J, Ye G, Shi K, et al. Structural basis of receptor recognition by Sars-Cov-2. Nature. 2020; 581 (7807), 221-4.
[17] Zhou P, Yang XL, Wang XG, et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature. 2020; 579 (7798), 270-3.
[18] Bartsch SM, Ferguson MC, McKinnell JA, et al. The potential health care costs and resource use associated with Covid-19 in the United States. Health Aff (Millwood). 2020; 39 (6), 927-35.
[19] Wong G, Bi YH, Wang QH, Chen XW, Zhang ZG, and Yao YG. Zoonotic origins of human coronavirus 2019 (Hcov-19 / Sars-Cov-2): why is this work important? Zool Res. 2020; 41 (3), 213-9.
[20] Rodriguez-Morales AJ, Dhama K, Sharun K, Tiwari R, and Bonilla-Aldana DK. Susceptibility of felids to coronaviruses. Vet Rec. 2020; 186 (17), e21.
[21] Tiwari R, Dhama K, Sharun K, et al. Covid-19: Animals, veterinary and zoonotic links. Vet Q. 2020; 40 (1), 169-82.
[22] Lam TT-Y, Shum MH-H, Zhu H-C, et al. Identifying Sars-Cov-2 Related Coronaviruses in Malayan pangolins. Nature 2020; 583 (7815), 282-5.
[23] Bonilla-Aldana DK, Cardona-Trujillo MC, Garcia-Barco A, et al. Mers-Cov and Sars-Cov Infections in animals: a systematic review and meta-analysis of prevalence studies. Infez Med. 2020; 28 (suppl 1), 71-83.
[24] Rabaan AA, Al-Ahmed SH, Haque S, et al. Sars-Cov-2, Sars-Cov, and Mers-Cov: a comparative overview. Infez Med. 2020; 28 (2), 174-84.
[25] Moher D, Liberati A, Tetzlaff J, Altman DG, and Group P. Preferred reporting items for systematic reviews and meta-analyses: the prisma statement. PLoS Med. 2009; 6 (7), e1000097.
[26] Institute of Health Economics (IHE). “Quality Appraisal of Case Series Studies Checklist.” Institute of Health Economics, http://www.ihe.ca/research-programs/rmd/cssqac/cssqac-about.
[27] Downes MJ, Brennan ML, Williams HC, and Dean RS. Development of a critical appraisal tool to assess the quality of cross-sectional studies (Axis). BMJ Open. 2016; 6 (12), e011458.
[28] Wallace BC, Dahabreh IJ, Trikalinos TA, Lau J, Trow P, and Schmid CH. Closing the gap between methodologists and end-users: r as a computational back-end. J Stat Softw. 2012; 49 (5).
[29] Viechtbauer W. Conducting meta-analyses in R with the metafor package. J Stat Soft. 2010; 36 (3).
[30] Kontopantelis E, and Reeves D. Performance of statistical methods for meta-analysis when true study effects are non-normally distributed: a comparison between dersimonian-laird and restricted maximum likelihood. Stat Methods Med Res. 2012; 21 (6), 657-9.
[31] Bonilla-Aldana DK, Jimenez-Diaz SD, Arango-Duque JS, et al. Bats in ecosystems and their wide spectrum of viral infectious potential threats: Sars-Cov-2 and other emerging viruses. Int J Infect Dis. 2021; 102, 87-96.
[32] Bonilla-Aldana DK, Jimenez-Diaz SD, Patel SK, et al. Importance of bats in wildlife: not just carriers of pandemic Sars-Cov-2 and other viruses. J. Pure Appl. Microbiol. 2020; 14 (Suppl. 1), 709-12.
[33] Bai Y, Urushadze L, Osikowicz L, et al. Molecular survey of bacterial zoonotic agents in bats from the country of Georgia (Caucasus). PLoS One. 2017; 12 (1), e0171175.
[34] Muscarella R, and Fleming TH. The role of frugivorous bats in tropical forest succession. Biol Rev Camb Philos Soc. 2007; 82 (4), 573-90.
[35] Riccucci M. Bats as Materia Medica: An ethnomedical review and implications for conservation. Vespertilio. 2013; 16, 249-70.
[36] Han HJ, Wen HL, Zhou CM, et al. Bats as reservoirs of severe emerging infectious diseases. Virus Res. 2015; 205, 1-6.
[37] Memish ZA, Mishra N, Olival KJ, et al. Middle East Respiratory Syndrome Coronavirus in bats, Saudi Arabia. Emerg Infect Dis. 2013; 19 (11), 1819-23.
[38] Monchatre-Leroy E, Boue F, Boucher JM, et al. Identification of alpha and beta coronavirus in wildlife species in France: bats, rodents, rabbits, and hedgehogs. Viruses. 2017; 9 (12), 364.
[39] Wang LF, and Anderson DE. Viruses in bats and potential spillover to animals and humans. Curr Opin Virol. 2019; 34, 79-89.
[40] Anindita PD, Sasaki M, Setiyono A, et al. Detection of coronavirus genomes in moluccan naked-backed fruit bats in Indonesia. Arch Virol. 2015; 160 (4), 1113-8.
[41] Hu D, Zhu C, Ai L, et al. Genomic characterization and infectivity of a Novel Sars-Like Coronavirus in Chinese bats. Emerg Microbes Infect. 2018; 7 (1), 154.
[42] Wang LF, and Eaton BT. Bats, civets and the emergence of Sars. Curr Top Microbiol Immunol. 2007; 315, 325-44.
[43] Sarkar J, and Guha R. Infectivity, virulence, pathogenicity, host-pathogen interactions of Sars and Sars-Cov-2 in experimental animals: a systematic review. Vet Res Commun. 2020; 44 (3-4), 101-10.
[44] Bonilla-Aldana DK, Quintero-Rada K, Montoya-Posada JP, et al. Sars-Cov, Mers-Cov and Now the 2019-Novel Cov: have we investigated enough about coronaviruses? - a bibliometric analysis. Travel Med Infect Dis. 2020; 33, 101566.
[45] Cascio A, Bosilkovski M, Rodriguez-Morales AJ, and Pappas G. The socio-ecology of zoonotic infections. Clin Microbiol Infect. 2011; 17 (3), 336-42.
[46] Kia GSN, Tao Y, Umoh JU, Kwaga JKP, and Tong S. Identification of Coronaviruses, Paramyxoviruses, Reoviruses, and Rotaviruses among bats in Nigeria. Am J Trop Med Hyg. 2021; 104 (3), 1106-10.
[47] Lu M, Wang X, Ye H, et al. Does public fear that bats spread Covid-19 jeopardize bat conservation? Biol Conserv. 2021; 254, 108952.
[48] Simsek C, Corman VM, Everling HU, et al. At least seven distinct rotavirus genotype constellations in bats with evidence of reassortment and zoonotic transmissions. mBio 2021; 12 (1), e02755-20.
[49] Wacharapluesadee S, Tan CW, Maneeorn P, et al. Evidence for Sars-Cov-2 related coronaviruses circulating in bats and pangolins in Southeast Asia. Nat Commun. 2021; 12 (1), 972.
[50] Irving AT, Ahn M, Goh G, Anderson DE, and Wang LF. Lessons from the host defences of bats, a unique viral reservoir. Nature. 2021; 589 (7842), 363-70.
[51] Luo J, Liang S, and Jin F. Gut microbiota in antiviral strategy from bats to humans: a missing link in Covid-19. Sci China Life. Sci 2021; 27, 1-15.
[52] do Vale B, Lopes AP, Fontes MDC, Silvestre M, Cardoso L, and Coelho AC. Bats, pangolins, minks and other animals - villains or victims of Sars-Cov-2? Vet Res Commun. 2021; 45 (1), 1-19.
[53] Hunter P. The spread of the Covid-19 Coronavirus: health agencies worldwide prepare for the seemingly inevitability of the Covid-19 Coronavirus Becoming Endemic. EMBO Rep 2020; 21 (4), e50334.
[54] Bonilla-Aldana DK, Holguin-Rivera Y, Perez-Vargas S, et al. Importance of the One Health Approach to Study the Sars-Cov-2 in Latin America. One Health. 2020; 10, 100147.
[55] Hemida MG, and Ba Abduallah MM. The Sars-Cov-2 Outbreak from a one health perspective. One Health. 2020; 10, 100127.
[56] Marty AM, and Jones MK. The novel coronavirus (Sars-Cov-2) Is a one health issue. One Health. 2020; 9, 100123.
[57] Simas PV, Barnabe AC, Duraes-Carvalho R, et al. Bat coronavirus in Brazil related to appalachian ridge and porcine epidemic diarrhea viruses. Emerg Infect Dis. 2015; 21 (4), 729-31.
[58] Brandao PE, Scheffer K, Villarreal LY, et al. A coronavirus detected in the vampire bat desmodus rotundus. Braz J Infect Dis. 2008; 12 (6), 466-8.
[59] Goes LGB, Campos ACA, Carvalho C, et al. Genetic Diversity of Bats Coronaviruses in the Atlantic Forest Hotspot Biome, Brazil. Infect Genet Evol. 2016; 44, 510-3.
[60] Goes LG, Ruvalcaba SG, Campos AA, et al. Novel Bat Coronaviruses, Brazil and Mexico. Emerg Infect Dis. 2013; 19 (10), 1711-3.
[61] Bonilla-Aldana DK, Suarez JA, Franco-Paredes C, et al. Brazil burning! What is the potential impact of the amazon wildfires on vector-borne and zoonotic emerging diseases? A statement from an international experts meeting. Travel Med Infect Dis. 2019; 31, 101474.
[62] Cibulski S, de Lima FES, and Roehe PM. Coronaviruses in Brazilian bats: a matter of concern? PLoS Negl Trop Dis 2020; 14 (10), e0008820.
[63] Dimkic I, Fira D, Janakiev T, et al. The Microbiome of bat guano: for what is this knowledge important? Appl Microbiol Biotechnol. 2021; 105 (4), 1407-19.